| |
| |
|
|
| |
|
|
Epidermal
growth factor receptor (EGFR) |
|
|
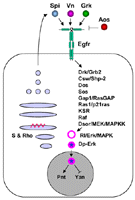
| Symbol: AKA: EGFR; Drosophila EGF receptor (DER) {Links}
|
Flybase ID: {Flybase_ID} |
| Synonyms: {Name}
|
{GadFly} |
| Function: {Short_Function} |
{LocusLink} |
| Keywords: {Keywords} |
{Interactive_Fly} |
{Summary}
|
|
|
- Vn/EGFR signaling directs cells to become notum by antagonizing wing
development and by activating notum-specifying genes (Wang,
2000)
- Vn/EGFR signaling directs cells to become part of the dorsal compartment
by induction of apterous and consequently also controls wing development,
which depends on an interaction between dorsal and ventral cells (Wang,
2000)
- Activated by small signalling molecules, all characterised by having
one EGF-like motif in their extracellular domain:
- Transforming growth factor a (TGF-a)
- EGF
- Neuregulins
|
|
|
- Required for vein initiation and subsequent differentiation
- Vn/EGFR signaling represses wing development
- Misexpressing constitutive receptor, EGFRltop4.2, in the presumptive
wing reduces the wing to a stump covered with sensilla characteristic
of the proximal wing (hinge) region and expression of the wing specific
gene vestigial was repressed (Wang,
2000)
- 4 ligans are known to modulate the function of DER
- Vein (vertebrate neuregulin isoform)
- Gurken (resemble TGF-a and are expressed as membrane-bound precursor
molecules)
- Spitz (resemble TGF-a and are expressed as membrane-bound precursor
molecules)
- Argos (novel inhibitory ligand)
- Double in-situ hybridization reveals that the medial region of EGFR
downregulation coincides with the region of kn gene expression
(Fig. 5B). In kn mutants, downregulation of EGFR expression
in the medial region is almost completely eliminated (Fig. 5C), indicating
that the downregulation of EGFR expression is dependent on kn. Reciprocially,
EGFR expression is greatly reduced in discs in which kn is ectopically
expressed under the control of the MS1096-Gal4 driver (Fig. 5D). (Mohler,
2000)
- vn is a target of EGFR signaling in the embryo (Golembo et
al. 1999; Wessells et al. 1999)
- Vn/EGFR signaling regulates Ap expression in a cell autonomous fashion
only early in wing development
- ap expression partially overlaps that of vn in the
second instar (Fig 4I) (Wang,
2000)
- ap can be induced ectopically in ventral clones misexpressing
an activated form of the receptor, EGFRlambdatop4.2
(Fig. 4J) (Wang,
2000)
- Egfrtsla mutant clones generated in the first
instar show autonomous loss of ap expression (Fig. 4K), whereas
clones generated in the second instar express ap normally
(data not shown) (Wang,
2000)
- loss of EGFR activity in whole discs from mid-first to mid-second
instar (with a temperature-sensitive allele) results in complete
loss of ap expression, whereas ap is still expressed
in discs from larvae given a temperature shift slightly later during
the second instar (Fig. 4L–N). (Wang,
2000)
|
|
|
| {Physical
interactions} |
Transcriptional
Regulation |
- Patterned activation in the embryonic epidermis is primarily modulated
by veinlet
- Normally depressed in the 3-4 intervein region in the wing imaginal
disc and is dependent on kn activity
|
|
|
| {Structure} |
Location (protein
and transcript) |
- Activated in the epidermis of the late embryo
- transcription is initially uniform throughout the wing blade, and
by the end of the 3rd instar EGFR transcription is repressed
along the wing margin and in a medial stripe across the wing blade region
(Guichard, 1999)
|
Protein
Modifications and Regulation |
| {Modifications} |
|
|
- In vertebrates, four different EGF receptor homologues constitute
the ErbB superfamily
|
|
|
- Inactivating Vn/EGFR activity (with temp sensitive alleles; Egfrts1a,
vntsWB240) during the second instar (a 24 hr period)
caused loss of the notum (Fig. 1 E,F) (Wang,
2000). The wing developed but showed pattern abnormalities characteristic
of vn hypomorphs (Fig 1E) (Wang,
2000). Later shifts during the third instar did not cause loss of
the notum (data not shown) (Wang,
2000). This demonstrates Vn/EGFR activity is required for notum
development in the second instar when wg is required to specify the
wing (Ng,
1996).
- In vn mutants, expression of Caup/Ara is lost
(Fig. 2E) and loss of EGFR signaling, in EGFRts clones,
in the medial notum resulted in a loss of Caup/Ara expression
(Fig. 2G). However, clones in the lateral notum continued to express
Caup/Ara (Fig. 2G), suggesting
other factors regulate Iro-C gene expression in these cells at this
stage (Wang,
2000)
|
Overexpression
/ Ectopic expression |
- ptc-GAL4 UAS-EGFR generates wings with fusion of veins 3 & 4 in
the proximal portion of the wing (Fig 5E) (Mohler,
2000), which is similar to weak fu or kn mutations,
suggesting that the downregulation oin
EGFR in the medial wing may play a role in the spacing of veins 3 &
4 (Mohler,
2000). More distally the spacing of veins 3 & 4 appears normal.
(Mohler,
2000)
- EGFR driven by the MS1096 driver does not cause formation of
ectopic vein between veins 3 & 4, although sgnificant amount of
ectopic vein material is induced anterior to vein 3 and posterior to
vein 4 (fig 5G) (Mohler,
2000) suggesting that the creation
of a vein-free zone between veins 3 & 4 is not likelly to be explained
solely by downregulation of EGFR by kn. (Mohler,
2000)
- Ectopic expression of an activated form of the receptor, EGFRlambdatop4.2
greatly reduced the size of the wing and partially transformed to hinge
as evident from groups of ectopic sensilla characteristic of the proximal
wing (hinge) and expression of vestigial was repressed cell autonomously
(Fig. 2C & 3A-D). (Wang,
2000).
|
|
|
| {Reagents} |
|
|
|
|
|