Carthew Lab: Research
-
RNA-Based Gene Regulation
-
-
Genomes contain thousands of genes and their expression is controlled by transcription. Although it was thought that transcription fidelity ensures that genes do not make double-stranded transcripts (dsRNA), this is not entirely true. Convergent transcription of neighboring genes can generate dsRNA where they overlap. In Drosophila, the 3‘UTRs of 2000 genes overlap in such a manner. Transcripts can also foldback into extended hairpins. Altogether, these dsRNAs generate endogenous siRNAs (endo-siRNAs) due to the action of the Dicer ribonuclease. As a result, the transcripts are cleaved and destroyed by Dicer. In several cases, the endo-siRNAs guide further silencing by association with an Argonaute protein. So perhaps hundreds or thousands of protein-coding genes are negatively regulated in this way.
-
-
How does this regulation work in the context of other regulatory mechanisms? It is not essential - mutation of the endo-siRNA Dicer or Argonaute in Drosophila does not lead to the animal’s premature death or disease (Lee et al., 2004 and Kim et al., 2007). What then do endo-siRNAs do? We know of one function for them during Drosophila embryogenesis. If an embryo is subjected to a temperature gradient over its body, the warmer regions will develop faster than the cooler regions but only up to a certain point. Thereafter, development compensates for the difference and the regions synchronize their development. This synchronization requires endo-siRNAs to be made (Lucchetta et al., 2009). If not, the embryo remains out of sync and dies. We speculate that endo-siRNAs are working like molecular thermostats, slowing gene expression down if it goes too fast because of higher temperature.
-
-
MicroRNAs
-
-
Certain transcripts can fold into short hairpin-loops that are a unique form of dsRNA. These hairpin-loops generate microRNAs (miRNAs) due to the action of Dicer. MicroRNAs associate with an Argonaute protein and unwind, retaining one RNA strand. However, miRNAs do not silence their own expression because of the unique way in which they are made. Their transcript precursors are first processed in the nucleus to make short hairpin-loops that prove resistant to silencing by the mature miRNA in the cytoplasm. Instead, a miRNA silences protein-coding genes whose transcripts bear complementary sequences in their 3‘UTRs. Basepairing does not need to be perfect for silencing; in fact, binding is typically imperfect between miRNA and transcript. Silencing occurs by transcript destabilization and inhibition of protein translation. How miRNAs inhibit translation is not clear and quite controversial.
-
-
We do not exactly know how many genes are regulated by a miRNA. Computational methods predict hundreds to thousands of genes are regulated by each miRNA because of the imperfect nature of miRNA binding. Systems biology experiments paint a more limited picture, predicting each miRNA binds and inhibits tens to hundreds of transcripts. Some of the genes that are silenced do very important things for the organism, and their silencing is paradoxically important to the organism as well. For example, one such silenced gene makes a protein that prevents germline stem cells from dividing in Drosophila. However, the stem cells produce miRNAs that silence the cell cycle inhibitor, thereby allowing the cells to divide and produce eggs (Hatfield et al., 2005 ). There are many examples of this kind of silencing, and many aspects of physiology and development depend on miRNAs.
-
-
MicroRNA genes arose at the dawn of animal evolution, and about 30 of these genes have remained conserved across the animal kingdom. The regions of the body where each of these genes is expressed has also changed very little across the animal kingdom. For example, the miRNA miR-7 is specifically expressed in neurosecretory cells of animals as diverse as annelid worms, insects, and mammals (Li et al., 2009 ). We still do not know whether these ancient miRNAs have conserved functions within the cell types in which they are expressed. MicroRNA evolution did not stop with the Cambrian explosion. It turns out that miRNA genes are the fastest evolving type of animal gene known. They are born at rates about ten-fold faster than protein-coding genes, and they seem to change very rapidly (Lu et al., 2008). At last count, there are 1400 miRNAs identified in humans and 240 miRNAs in Drosophila. Clearly, they are a rapidly growing gene family.
A Systems Biology View of miRNAs
A paradox for many miRNAs is that they lack strong phenotypic consequences when mutated and yet they are evolutionarily conserved. Some have argued that this paradox is due to the weak repression of target gene expression elicited by most miRNAs. However, miRNAs also exist in higher-order relationships, and less emphasis has been placed on the biochemical networks that include miRNAs. Yet this is fundamental to understand what roles miRNAs play. Most biochemical networks are robust; things happen reproducibly and uniformly even in the face of variability that can be induced by the environment, genetic variation, and random chance. Biological processes, particularly irreversible ones such as differentiation, are strongly robust to ensure a minimal impact of error. We think that miRNAs help to generate the robustness of biochemical networks.
Genome-wide experiments and network modeling support a link between miRNAs and robustness. Focused experimentation on individual miRNAs corroborate the genome-wide studies. In Drosophila, the transcription repressor YAN binds and represses the miR-7 gene. In turn, miR-7 represses protein expression of YAN. YAN and miR-7 are part of a network that regulates the transition of multipotent progenitor cells to become differentiated photoreceptors (Li and Carthew, 2005). The network is a bistable system that can transition from a high YAN/low miR-7 state to a high miR-7/low YAN state. These two stable states dictate whether a cell remains multipotent or differentiated, respectively. EGF Receptor signaling induces switching from the multipotent to differentiated state.
YAN and miR-7 act within several feedback and feedforward loops in the network. The role of these loops is to generate robustness to the network. When development is perturbed in a miR-7 mutant by oscillating the growth temperature, the network switches states less robustly, and errors in cell differentiation are observed (Li et al., 2009 ). These errors are undetectable under uniform temperature conditions. If miR-7 is not mutated, the network is robust under either uniform or oscillating temperature growth conditions. This miRNA makes differentiation robust to mild environmental perturbation.
Disease and MicroRNAs
Given the sheer number of miRNAs and their fundamental function in gene regulation, it is not surprising to find them linked to or causative of many human diseases. These are:
-
•Cancer - leukemia, liver, lung, glioma, renal, pancreatic, colon, breast, prostate
-
•Cardiac hypertrophy and ischemia
-
•Muscular dystrophy
-
•Lou Gehrig’s Disease
-
•Alzheimers Disease
-
•Parkinsons Disease
-
•Tourettes Syndrome
-
•Psoriasis
-
•Diabetic nephropathy
Anti-miRNA oligonucleotides show promise as a new class of therapeutic treatment for some of these diseases.
The Future
Our objective is to understand the principles of gene regulation by endo-siRNAs and miRNAs in animals as a means to understand and treat human disease. One research aim is to use systems-level approaches to gain a complete picture about how miR-7 provides robustness to the YAN network. This aim utilizes gene recombineering, live-time-lapse imaging of fluorescently tagged proteins, next-generation sequencing, and quantitative computer modeling, with the help of Luis Amaral’s group at Northwestern. Another aim is to explore the role of miRNAs in adaptation during selection. Robustness operates on the evolutionary scale as well, and miRNAs might inhibit adaptation when populations are selected for certain traits. We are using artificial selection paradigms coupled with next-generation sequencing. A third aim is to determine whether a miRNA elicits robustness by regulating a handful of target genes or by its overall effect on a dynamic transcriptome. An intriguing idea is that the level of miRNA regulation for any given transcript depends on its transcriptome environment - if other transcripts that bind the miRNA are more abundant, then less miRNA is available to regulate the transcript. A shifting transcriptome responding to a variable could be buffered by a counteracting shift in miRNA partitioning between transcripts.
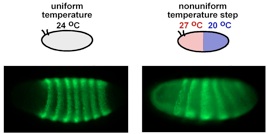
Embryos lacking the Ago2 gene were incubated in a microfluidics chamber that controls the environmental temperature. Left, an embryo incubated in a uniform bathing temperature of 24C and right, an embryo incubated in a bath where the anterior (head) half was warmer than the posterior (tail) half. Confocal micrographs show protein expression of the segmental patterning gene Eve in green. Normally there are seven uniform stripes of Eve expression, but this is disrupted in the embryo to the right.
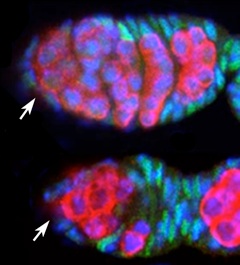
Confocal laser micrograph of two Drosophila ovaries. Female germ cells are marked in red, support follicle cells are marked in green, and cell nuclei in blue. The top ovary is from a normal animal and the bottom ovary from an animal missing all of its miRNAs. The arrows point to the germline stem cells. Fewer germ cells are present in the bottom ovary because the stem cells are dividing slowly.
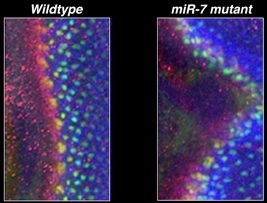
Confocal laser micrograph of two Drosophila eyes taken from young animals growing in a oscillating temperature environment. Micrograph shows protein expression of the network genes ATONAL (red), SENSELESS (green), and YAN (blue). The left eye is from an animal with normal copies of its miR-7 gene, while the right eye is from an animal with mutant copies of miR-7. Gene expression is abnormal in the eye on the right.

Northwestern University | Northwestern Search
Center for Cell and Developmental Systems Biology | Department of Molecular Biosciences
2205 Tech Drive, Hogan 2-100, Evanston, IL 60208 Email the Webmaster
WWW Disclaimer and University Policy Statements © 2011 Northwestern University